
Effect and mechanism of Xuanfu Daizhe Decoction on proliferation, migration and invasion activities of esophageal cancer cells
-
摘要:目的
探讨旋覆代赭汤对食管癌细胞增殖、迁移和侵袭活性的影响及机制。
方法使用不同质量浓度的旋覆代赭汤处理人食管癌细胞系EC109, 并依据旋覆代赭汤的质量浓度将细胞分为高浓度组(200 μg/mL)、中浓度组(100 μg/mL)、低浓度组(50 μg/mL)和空白组(0 μg/mL)。分别采用CCK-8实验、划痕实验和Transwell侵袭实验检测各组细胞的增殖、迁移和侵袭活性。采用蛋白质印迹法(Western blot)和细胞免疫荧光染色法检测各组细胞中糖酵解相关酶[己糖激酶2(HK2)、乳酸脱氢酶A(LDHA)、磷酸果糖激酶1(PFK1)]的蛋白表达水平, 并采用实时荧光定量聚合酶链式反应(qRT-PCR)检测上述酶编码基因mRNA相对表达量。构建裸鼠负瘤模型,分为对照组和旋覆代赭汤饲喂组(20 mg/kg), 观察旋覆代赭汤对食管癌体内生长的影响。
结果CCK-8实验、划痕实验和Transwell侵袭实验结果显示,旋覆代赭汤可浓度依赖性抑制EC109细胞的增殖、迁移和侵袭活性。Western blot和细胞免疫荧光分析结果显示,与空白组相比,低浓度组、中浓度组、高浓度组细胞的HK2、LDHA、PFK1蛋白表达水平降低,差异有统计学意义(P < 0.05); qRT-PCR检测结果显示,与空白组相比,低浓度组、中浓度组、高浓度组细胞的HK2 mRNA、LDHA mRNA、PFK1 mRNA相对表达量均降低,差异有统计学意义(P < 0.01或P < 0.000 1)。覆代赭汤饲喂组裸鼠背部皮下的肿瘤体积小于对照组,肿瘤组织中LDHA蛋白表达水平低于对照组,促凋亡蛋白Bax表达水平高于对照组,差异有统计学意义(P < 0.05)。
结论旋覆代赭汤可浓度依赖性抑制食管癌细胞糖酵解过程,进而抑制细胞增殖、迁移、侵袭及体内肿瘤生长。
Abstract:ObjectiveTo investigate the effect and mechanism of Xuanfu Daizhe Decoction on the proliferation, migration, and invasion activities of esophageal cancer cells.
MethodsHuman esophageal cancer cell line EC109 was treated with Xuanfu Daizhe Decoction at different concentrations, and the cells were divided into high-concentration group (200 μg/mL), medium-concentration group (100 μg/mL), low-concentration group (50 μg/mL), and blank group (0 μg/mL) based on the concentration. CCK-8 assay, wound healing assay, and Transwell invasion assay were used to detect the proliferation, migration, and invasion activities of the cells in each group, respectively. Western blot and cellular immunofluorescence staining were employed to detect the protein expression levels of glycolysis-related enzymes [hexokinase 2 (HK2), lactate dehydrogenase A (LDHA), and phosphofructokinase 1 (PFK1)] in the cells of each group, and real-time quantitative fluorescent polymerase chain reaction (qRT-PCR) was used to detect the relative expression levels of mRNA encoding these enzymes. A nude mouse tumor-bearing model was established and divided into control group and Xuanfu Daizhe Decoction-fed group (20 mg/kg) to observe the effect of Xuanfu Daizhe Decoction on the growth of esophageal cancer in vivo.
ResultsThe results of CCK-8 assay, wound healing assay, and Transwell invasion assay showed that Xuanfu Daizhe Decoction could inhibit the proliferation, migration, and invasion activities of EC109 cells in a concentration-dependent manner. Western blot and cellular immunofluorescence analysis revealed that compared with the blank group, the protein expression levels of HK2, LDHA, and PFK1 in the low-concentration, medium-concentration, and high-concentration groups were decreased (P < 0.05). The qRT-PCR results showed that compared with the blank group, the relative expression levels of HK2 mRNA, LDHA mRNA, and PFK1 mRNA in the low-concentration, medium-concentration, and high-concentration groups were all reduced(P < 0.01 or P < 0.000 1). The tumor volume in the subcutaneous tissue on the back of nude mice in the Xuanfu Daizhe Decoction-fed group was smaller than that in the control group, and the protein expression level of LDHA in the tumor tissue of the Xuanfu Daizhe Decoction-fed group was lower than that in the control group, while the expression level of the pro-apoptotic protein Bax was higher than that in the control group (P < 0.05).
ConclusionXuanfu Daizhe Decoction can inhibit the glycolysis process of esophageal cancer cells in a concentration-dependent manner, thereby inhibiting cell proliferation, migration, invasion, and tumor growth in vivo.
-
Keywords:
- Xuanfu Daizhe Decoction /
- esophageal cancer /
- glycolysis /
- proliferation /
- migration /
- invasion
-
视神经脊髓炎谱系疾病(NMOSD)是一种由体液免疫、细胞免疫参与,累及视神经、脊髓的中枢神经系统(CNS)炎性脱髓鞘性疾病[1],其发病率与种族、地理位置具有相关性。流行病学研究[2]显示,非高加索人群对NMOSD易感性高,NMOSD可见于各年龄阶段,但以青壮年多见,成人水通道蛋白4(AQP4)抗体阳性亚群中以女性为多。2015年国际视神经脊髓炎(NMO)诊断小组更新了诊断标准[3],并统一使用“视神经脊髓炎谱系疾病”这一术语。大多数NMOSD患者血清或脑脊液中可发现AQP4抗体阳性[4], NMOSD发病除由抗体介导外,还有多种免疫细胞的参与,包括B细胞、T细胞,此外细胞因子及肠道菌群也在其中发挥着重要作用。本文总结了NMOSD发病机制的研究进展,现综述如下。
1. AQP4抗体
AQP4是水通道蛋白家族的一员,能够快速转运水的膜蛋白,在星形胶质细胞、室管膜细胞中表达,发挥调控脑组织水稳态的作用。然而AQP4在胃肠道、肾脏等外周器官中也会表达,因外周存在补体调节蛋白,可避免表达AQP4的外周组织受到损伤[5]。AQP4抗体属于免疫球蛋白G(IgG)1亚类,最常见的亚型是M1、M23,M1、M23亚型以及不同区域和器官中补体调节蛋白表达存在差异,故表达AQP4组织对NMOSD易感性也有差异[6-7]。CNS细胞中可见M23表达明显升高,随着M23丰度的增加,M1-AQP4完全内化促进正交粒子阵列(OAP)形成[8],OAP是补体C1q与AQP4抗体结合的关键。
60%~70%的NMOSD患者AQP4抗体呈阳性[2], AQP4抗体是NMOSD的特异性标志物。人源性AQP4抗体在体内、体外均可诱导NMOSD病理性改变。AQP4抗体与星形胶质细胞AQP4结合引起神经系统炎症急性发作,并诱发CNS出现相应病理改变,包括星形胶质细胞病变、补体依赖性细胞毒性、抗体依赖性细胞毒性、血脑屏障(BBB)破坏、神经炎症、脱髓鞘和神经元损伤。
细胞外环C是人类AQP4抗体最常见的靶点[9], 在易感人群中,反应性T细胞、抗原抗体反应有利于细胞外环C暴露,促进幼稚T细胞呈递白细胞介素-6膜结合受体(MIL-6R)/白细胞介素(IL)-6B从而使IL-6水平升高,促进辅助性T细胞(Th)17细胞分化进一步刺激B细胞激活,诱导产生AQP4抗体介导NMOSD发生。一项纵向研究[10]发现,AQP4抗体滴度在复发时升高3倍,经免疫治疗后抗体滴度降低,提示AQP4抗体滴度与疾病的活动性存在相关性[11]。另一项研究[12]表明,抗体滴度的升高可提示疾病发作期的严重程度,提示AQP4抗体参与NMOSD的发生。
2. 补体途径
补体系统在固有免疫监控中发挥着至关重要的作用,包括启动对病原体的防御反应以及清除有毒有害物质来维持宿主的动态平衡。AQP4抗体与抗原结合后激活补体,可引起原发性星形胶质细胞损伤和炎症反应,同时造成脱髓鞘和神经元损伤,所以抑制补体激活可减轻抗原抗体结合后的下游反应。
在NMOSD患者中,AQP4抗体可与视神经、脊髓上AQP4的表位结合导致反应性星形胶质细胞生成、粒细胞募集,从而激活补体级联反应。AQP4抗体通过经典途径导致补体激活产生C3a、C5a, 表明补体途径参与NMOSD的发生。一项实验[13]表明, C3a在CNS过量表达后可加重实验性自身免疫性脑脊髓炎(EAE)小鼠血管周围炎性细胞的浸润。补体成分C5a既是促炎因子,也是中性粒细胞、嗜酸性粒细胞的趋化因子,可破坏BBB功能,刺激补体激活[9], 从而导致炎症反应扩大、AQP4内化、兴奋性氨基酸转运体2下调,并导致谷氨酸兴奋性毒性以及补体依赖、抗体依赖的细胞毒性。补体成分C5b可促进膜攻击复合物形成从而产生细胞毒性。以上研究均表明,补体途径在NMOSD发病中发挥作用。补体C5抑制剂Eculizumab是一种新型的人源化IgG2/4单克隆抗体,可选择性地与补体成分C5高亲和力地结合抑制其裂解成C5a和C5b。一项开放性试验[14]证实了C5抑制剂的有效性,另一项关于补体抑制剂Eculizumab的随机双盲Ⅲ期临床试验[15]发现,长期应用Eculizumab可持续降低AQP4阳性NMOSD复发风险,该药于2019年已在美国等国家被批准用于成人AQP4抗体阳性NMOSD患者的治疗。目前,针对补体途径不同组成部分药物的临床试验正在进行中[16],期待未来有更多有效的药物可用于NMOSD的临床治疗。
3. B细胞
B细胞通过分泌AQP4抗体、呈递抗原、产生细胞因子等多方面影响NMOSD的发生。脑脊液中AQP4抗体滴度相对较低甚至不存在,故认为B细胞首先在外周被激活并合成AQP4抗体[17], AQP4抗体通过BBB通透性增强的区域进入CNS。EAE模型中B细胞可以分泌IL-6, IL-6可进一步促进B细胞产生AQP4抗体[18]。在NMOSD中,B细胞首先在CNS外被激活发生体液免疫反应,同时B细胞通过BBB交换使血液、脑脊液中存在抗体相关B细胞和浆细胞[19]。
细胞表面CD19、CD20的表达可用于识别B细胞,CD20在B细胞上表达,但在浆细胞上显著下调[20],而CD19在前B细胞、浆细胞和浆母细胞上均表达。这些细胞表面蛋白是B细胞靶向治疗的常见靶点,如CD20单克隆抗体Rituximab可降低疾病复发率,提示B细胞耗竭治疗有效,但Rituximab治疗并非对所有患者有效,因外周浆细胞、浆母细胞不表达CD20, 产生AQP4抗体的细胞没有耗尽[21]。抗CD19单克隆抗体Inebilizumab能够直接杀伤产生抗体的浆母细胞、浆细胞,其抑制抗体的能力比Rituximab更全面。Inebilizumab的Ⅲ期临床试验[22]中,随机对照试验显示,应用Inebilizumab后扩展残疾状态量表(EDSS)评分恶化人数减少,改良Rankin评分量表(MRS)评分有更高的改善可能性,急性期受累病灶数量减少,住院次数减少,且CD20+细胞持续减少, N-MOmentum实验显示,长期应用Inebilizumab治疗能持续减少NMOSD复发次数,延缓残疾的发生。
3.1 记忆性B细胞
研究[23]发现,采用Natalizumab治疗后, NMOSD患者外周血B细胞由调节性B细胞转换为记忆B细胞表型,证实记忆B细胞增多可加重疾病。记忆B细胞分泌IL-6可能通过促进浆细胞存活、刺激AQP4-Ab产生、破坏BBB完整性、促进Th17分化等不同机制增强疾病活动性。
3.2 浆细胞
NMOSD患者外周血产生AQP4抗体的浆细胞数量显著增加,复发时浆细胞数目可达到高峰[24], 而脑脊液中几乎检测不到浆细胞[25]。NMOSD外周血中具有浆细胞表型的CD19intCD27highCD38highCD180-B细胞(PBs)可见明显扩增,在IL-6刺激后分泌AQP4抗体。在NMOSD中,升高的PBs以CD138+人类白细胞抗原-DR(HLA-DR)+表型为主,HLA-DR+PB表型频率增加表明抗原驱动B细胞活化。有学者[26]在脑脊液中发现B细胞具有体细胞超突变信号,存在抗原识别,同时在脑脊液中发现B细胞活化因子(BAFF)、增殖诱导配体(APRIL)、趋化因子CXC配体13(CXCL13)和IL-6水平升高, PBs在产生AQP4-Ab的细胞募集和维持中起关键作用。
3.3 调节性B细胞
调节性B细胞(Breg)可通过多种途径影响T细胞活性及细胞因子的表达。在NMOSD患者外周血中, Breg绝对数量减少[27]。使用类固醇或免疫抑制剂的系统性红斑狼疮患者中,CD19+CD24highCD38high Breg数量增加,研究[28]同时发现调节性B细胞中CD19+CD39+CD1d+IL-10+(B10)在NMOSD患者中细胞频率升高, B10细胞与AQP4抗体滴度存在相关性, B10细胞对AQP4抗体介导的炎症级联反应可出现代偿性增殖。
3.4 BAFF
BAFF是肿瘤坏死因子家族中的一员,由单核细胞、树突状细胞、骨髓基质细胞等多种类型细胞产生。BAFF受体与B细胞结合时激活经典、非经典核因子-кB(NF-кB)信号通路。NMOSD患者血清及脑脊液中BAFF、增殖诱导配体(APRIL)水平显著升高,BAFF水平与外周血B细胞的数量和BAFF受体的表达呈负相关[29]。血清BAFF水平升高与自身免疫性疾病抗体水平升高有关[30]。CNS星形胶质细胞可产生BAFF,在NMOSD患者中,血清BAFF水平升高可致过渡性B细胞增多,促进AQP4抗体生成,进一步促进B细胞和T细胞反应[31]。
4. T细胞
NMOSD是一种致残甚至致命的CNS自身免疫性炎症性脱髓鞘疾病。动物试验[32]表明,效应性T细胞可能在AQP4抗体阳性NMOSD小鼠的发病机制中发挥作用。在NMOSD患者中, AQP4特异性T细胞出现明显扩增并表现出Th17极化,且Th17细胞数量以及Th17细胞因子IL-17a、IL-6、IL-21水平在脑脊液和血清中均增加或升高。AQP4抗体与髓鞘特异性T细胞共同转移至野生型小鼠可诱导CNS炎症病变[33]。由此表明,T细胞在NMOSD发病机制中具有潜在作用。
4.1 Th17细胞
Th17细胞的标志性细胞因子是IL-17家族,由幼稚CD4+ T细胞分化而来。T细胞受体(TCR)识别抗原呈递细胞呈递的抗原,由IL-6、转化生长因子-β(TGF-β)通过信号转导及转录激活因子3、Smads共同刺激,以激活Th17细胞特异性转录因子类视黄醇相关受体Yt磷酸化启动Th17分化,进一步促进B细胞分化和AQP4抗体产生。
NMOSD急性发作期, Th17细胞水平升高,IL-17+调节性T细胞(Treg)在NMOSD缓解期的细胞比例高于急性发作期,提示该细胞在控制疾病状态中起重要作用[28]。NMOSD患者脑脊液中IL-17A、IL-8水平与疾病总脊髓病变程度呈正相关[34], 表明NMOSD患者存在明显Th17相关反应,经治疗后炎症减轻,视神经结构完整性得以保留,Th17细胞及其相关细胞因子通过多种途径参与NMOSD疾病的CNS炎症反应。
NMOSD急性期患者经免疫治疗后记忆Th17细胞减少; 一项转移致病性AQP4反应性T细胞至野生型小鼠的试验[35]发现, AQP4反应性T细胞(尤其是Th17细胞)能够触发针对视神经、脊髓的炎症。
4.2 Treg
Treg在免疫反应中扮演重要的角色,当Treg频率或功能降低时可导致自身免疫性疾病。NMOSD患者Treg中, TNF受体相关因子1(TRAF1)、人类白细胞抗原-DRB1(HLA-DRB1)表达显著降低, TRAF1可促进NF-кB信号通路激活促进外周Treg存活。NMOSD患者外周血单个核细胞(PBMC)中叉头样转录因子3(Foxp3)显著降低[36], Foxp3在静息、活化的Treg中表达显著升高[37], 表明Treg参与NMOSD的发生。
4.3 滤泡辅助性T细胞(Tfh)
Tfh数量减少或功能失调可导致自身免疫性疾病。Tfh可分为Tfh2、Tfh17、Tfh1,Tfh2+Tfh17/Tfh1升高与NMOSD疾病活动性相关[38]。一项研究[39]发现,NMOSD患者CC趋化因子受体7(CCR7)-、CCR7-可诱导共刺激分子(ICOS)+记忆Tfh细胞比例与血浆中AQP4抗体水平呈正相关,CCR7+和CCR7+ICOS+记忆Tfh细胞百分比与脑脊液白细胞计数、脑脊液蛋白水平和脑脊液IL-21水平均呈正相关,提示记忆Tfh细胞参与外周AQP4抗体产生的同时还参与NMOSD的CNS免疫反应。
5. 固有淋巴细胞(ILCs)
研究[40]表明,在NMOSD患者外周血淋巴细胞亚群中,急性发作期和缓解期外周血CD19+ B细胞水平显著高于健康人群,且与脊髓病变长度呈正相关, CD4+/CD8+细胞比率和自然杀伤(NK)细胞(CD56+CD16)水平显著低于健康人群,外周血淋巴细胞亚群水平与疾病状态有关。
2型固有淋巴细胞(ILC2)是缺乏克隆分布抗原受体的特异性先天效应细胞,可产生2型免疫相关细胞因子,其作用是调节免疫和炎症反应。ILC2可与警示信号(如IL-33)相互作用产生IL-5、IL-13, 导致外周组织启动2型免疫反应[41]。一项研究[42]发现, NMOSD患者外周血中ILC2水平降低,而在颅内注射人NMOSD IgG和补体诱导的NMOSD小鼠模型中,小鼠大脑中表达IL-5的ILC2浸润明显增加,应用ILC2拮抗剂可导致CNS损伤体积增大、CNS中糖代谢降低、星形胶质细胞损伤和脱髓鞘加重,同时补体末端通路活性标志物C5b-9增加,表明ILC2缺失可使NMOSD进一步恶化,而ILC2扩增后星形胶质细胞损伤减少,提示ILC2可作为NMOSD潜在的治疗靶点。在EAE模型中, IL-33激活CNS中ILC2通过减弱Th17反应来改善疾病[43]。
6. 细胞因子
NMOSD患者血清、脑脊液中IL-6水平升高,急性期尤其显著, IL-6可与IL-6受体结合,通过不同途径广泛参与NMOSD的病理生理过程,如浆细胞诱导T细胞/B细胞异常活化,产生Th17细胞、AQP4抗体。
IL-6由多种类型细胞产生。NMOSD急性期脑脊液中IL-6、可溶性IL-6受体水平增高[23]。IL-6与B细胞相互作用促进炎症进展,诱导B细胞分化成浆细胞,维持外周浆细胞存活及浆母细胞分泌AQP4抗体[44],同时诱导T细胞活化促进Th17生成。
机体发生感染时, IL-6迅速合成,诱导急性期蛋白促进宿主防御和炎症反应。相关研究[45]显示, IL-6可加重小鼠抗体依赖性脊髓损伤;另一项研究[46]提出, IL-6与脊髓病灶长度、脑脊液AQP4抗体滴度、炎性细胞计数具有相关性。对于难治性NMOSD, IL-6受体拮抗剂tocilizumab可阻断IL-6与受体结合,同时减弱B细胞产生抗体能力,显著降低复发率,延长复发时间,提示tocilizumab可能对耐药的NMOSD患者有效[47]。人源化单克隆抗体satralizumab的临床试验[48]显示, satralizumab可显著降低AQP4阳性NMOSD复发风险。
粒细胞-巨噬细胞集落刺激因子(GM-CSF)由T细胞产生,协同IL-23、IL-1诱导巨噬细胞、树突细胞分泌IL-23、IL-6。NMOSD病变小鼠应用GM-CSF后可增加病变风险[49], GM-CSF在致病性Th17细胞扩增中起重要作用。
IL-17在NMOSD患者血清、脑脊液中水平升高,且复发期IL-17升高尤为明显,加重NMOSD患者病变部位炎性渗出、组织坏死,故复发期升高的IL-17可解释活动性NMOSD病变中中性粒细胞的高浓度。IL-17基因多态性与AQP4抗体阳性疾病相关,提示IL-17在NMOSD疾病发展中起重要作用,能促进抗体产生和炎症细胞浸润。
NMOSD中升高的细胞因子还有IL-21,IL-21是一种多向性细胞因子,可在体外与CD40L结合促进Th17增殖并加重炎症反应,抑制保护性Treg分化,进一步诱导B细胞向浆细胞分化。此外, IL-21的信号转导可促进Th17分化并刺激IL-23受体表达, IL-21通过促进Th17分化加重NMOSD炎症反应。
7. 肠道菌群
肠道菌群是人体中最大的微生物库,对维持宿主体内免疫和神经系统平衡至关重要。NMOSD患者的细胞经大肠杆菌刺激后表现出更显著的Th17反应性,与IL-1、IL-6、IL-17增多和IL-10减少有关[50]。NMOSD患者外周血单个核细胞培养中,CD4+ T细胞增殖更快, IL-6、IL-17水平升高,而这些改变与神经功能障碍、脂多糖水平均相关[50],脂多糖可能通过激活免疫系统破坏BBB。肠道菌群的代谢产物短链脂肪酸(SCFAs)可以直接调节机体免疫反应,抑制炎症反应。NMOSD患者粪便中SCFAs水平显著降低,而且粪便中乙酸和丁酸与疾病严重度呈负相关,提示其可能在NMOSD的发生中起重要作用[51]。SCFAs促进Treg数目增加,抑制Th17分化,同时影响BBB的发育和维持。Treg有利于髓鞘再生、少突胶质细胞分化而起到神经保护作用。无菌(GF)小鼠Th17细胞数量减少,分段丝状细菌定植GF小鼠可见Th17细胞增殖,肠道共生梭菌属改变可影响小鼠Th17和Treg之间的平衡[52]。
NMOSD患者T细胞出现增殖,主要表现为Th17表型,同时T细胞与产气荚膜梭菌(CPs)蛋白中的同源肽序列发生交叉反应, CPs促进Th17产生且可对AQP4产生免疫反应[53]。NMOSD患者肠道微生物群中CPs明显富集[54], CPs过度生长引起AQP4特异性T细胞和B细胞的增殖,促进NMO的发生与发展。CPs中B型、D型可产生具有神经毒性的ε毒素(ETX)损伤神经元、星形胶质细胞及少突胶质细胞[55]。CPs与肠道M蛋白结合,同时高度特化吞噬微生物抗原并将其呈递给抗原提呈细胞(APCs),经过APCs处理后的CPs可能引起AQP4特异性T细胞和B细胞反应,促使浆细胞生成增多,进而促进NMOSD发生。
8. 其他
高浓度氯化钠、高盐膳食摄入能促进Th17分化,导致严重EAE,因IL-23信号下游分子血清糖皮质激素激酶1在盐浓度升高后增加,促进IL-23受体表达,增强Th17分化,一项前瞻性研究[52]表明较高的钠摄入量与多发性硬化(MS)疾病活动度增加呈正相关。
在缺氧条件下,丝裂原活化蛋白激酶p38、NF-кB磷酸化增加向小胶质细胞诱导星形胶质细胞AQP4过表达,体温过低会增加星形胶质细胞质膜中AQP4丰度。
维生素D可诱导细胞的激活、增殖、凋亡和T细胞分化、浆细胞产生、基因表达以及细胞因子、抗体产生,维生素D水平降低与NMOSD的疾病活动状态相关,其参与NMOSD发病过程[56]。一些研究表明,血清25-羟基维生素D水平与疾病严重程度、进展、治疗效果均具有相关性。
妊娠晚期、产后期雌激素水平升高,患者的NMOSD复发率增高,此外NMOSD孕妇出现流产、先兆子痫等并发症的概率高,而产后6个月左右并发症可反向促进NMOSD复发[57]。小鼠试验[58]结果表明, AQP4抗体与孕妇胎盘中的合胞滋养层结合后通过激活补体方式潜在导致流产率增加,提示妊娠后激素水平与NMOSD存在相关性。
9. 总结与展望
NMOSD是一组以自身免疫异常引起的脑、脊髓和视神经髓鞘损伤为特征的疾病,随着AQP4抗体等诊断标志物的出现,临床人员对NMOSD有了更深入的了解。外周B细胞可产生AQP4抗体,该抗体可与抗原结合触发补体级联反应,形成膜攻击复合物、抗体及补体依赖性细胞毒性。AQP4抗原刺激可致Th17免疫反应及Th17相关细胞因子如IL-6的分泌, IL-6又可促进AQP4产生、T细胞向Th17分化,从而刺激B细胞激活,同时增强BBB的通透性,促进抗体和促炎细胞向CNS浸润,故细胞免疫、抗体、补体途径等均参与NMOSD的发生,但具体的细胞免疫表型的致病机制还需继续深入了解。肠道中CPs通过破坏BBB完整性或Th17与Treg之间平衡参与NMOSD发病机制,未来还需进一步研究肠道菌群的改变是否引起相应细胞亚群、细胞免疫表型、细胞因子的变化,是否可通过饮食干预、药物治疗、粪便微生物群移植等方法改变NMOSD患者肠道菌群而达到治疗疾病的目的,以及是否可通过调节相应细胞亚群、细胞因子等方式减轻NMOSD炎症反应,以期找到新的治疗靶点。
-
![]()
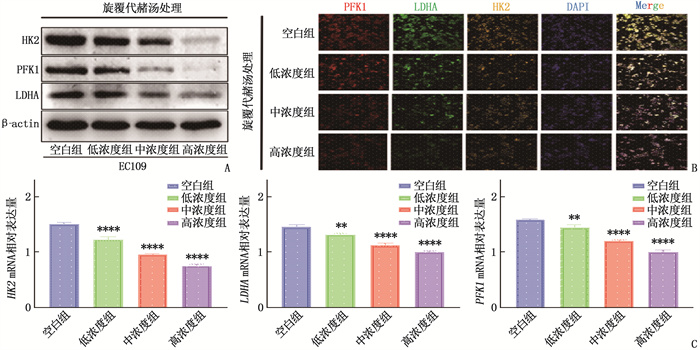
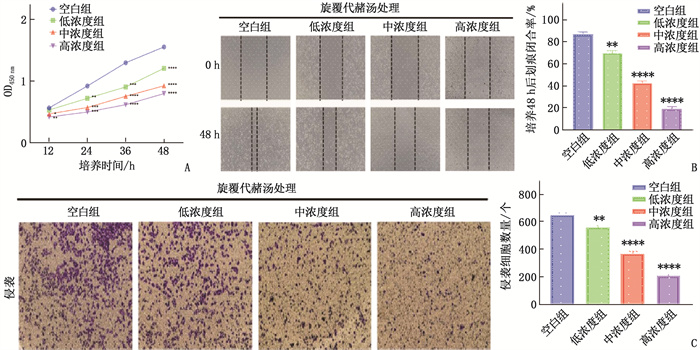
图 1 不同质量浓度旋覆代赭汤对EC109细胞增殖、迁移和侵袭活性的影响
A: 各组EC109细胞增殖活性的CCK-8实验检测结果; B: 各组EC109细胞迁移活性的划痕实验检测结果; C: 各组EC109细胞侵袭活性的Transwell侵袭实验检测结果。与空白组比较, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.000 1。
![]()
图 2 不同质量浓度旋覆代赭汤处理对食管癌细胞糖酵解的影响
A: 各组EC109细胞糖酵解相关酶蛋白表达水平的Western blot检测结果; B: 各组EC109细胞糖酵解相关酶蛋白表达水平的细胞免疫荧光检测结果; C: 各组EC109细胞糖酵解相关酶编码基因mRNA相对表达量的qRT-PCR检测结果(与空白组比较, **P < 0.01, ****P < 0.000 1)。
![]()
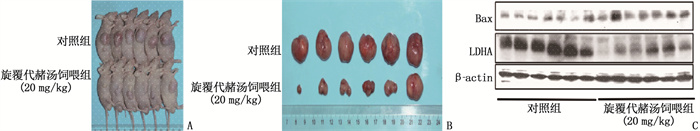
图 3 旋覆代赭汤饲喂对负瘤模型裸鼠肿瘤生长的影响
A、B: 2组负瘤模型裸鼠肿瘤生长情况比较; C: 2组裸鼠肿瘤组织中糖酵解和凋亡相关蛋白表达水平的Western blot检测结果。
表 1 引物序列
基因 引物序列(5′→3′) GAPDH 正向: TGTGGGCATCAATGGATTTGG 反向: ACACCATGTATTCCGGGTCAAT HK2 正向: AATGGACAACTGGTCGTGGAC 反向: CCCTCCAGGGGATCTGTTTG LDHA 正向: CTCACCGGATGCACCAATGTT 反向: CGCGTTGCTCACAATGTTCAT PFK1 正向: GGAGCGAGATCCCTCCAAAAT 反向: GGCTGTTGTCATACTTCTCATGG  下载: 导出CSV
下载: 导出CSV
-
[1] BÖHME F, RACZ K, SEBESTA C J, et al. Das ösophaguskarzinom[J]. Wien Med Wochenschr, 2023, 173(9): 209-215.
[2] ZHU H, MA X, YE T, et al. Esophageal cancer in China: Practice and research in the new era[J]. Int J Cancer, 2023, 152(9): 1741-1751. doi: 10.1002/ijc.34301
[3] LUENGO A, GUI D Y, VANDER HEIDEN M G. Targeting metabolism for cancer therapy[J]. Cell Chem Biol, 2017, 24(9): 1161-1180. doi: 10.1016/j.chembiol.2017.08.028
[4] BARBA I, CARRILLO-BOSCH L, SEOANE J. Targeting the Warburg effect in cancer: where do we stand[J]. Int J Mol Sci, 2024, 25(6): 3142. doi: 10.3390/ijms25063142
[5] CHELAKKOT C, CHELAKKOT V S, SHIN Y, et al. Modulating glycolysis to improve cancer therapy[J]. Int J Mol Sci, 2023, 24(3): 2606. doi: 10.3390/ijms24032606
[6] ZHENG S, LI H, LI Y, et al. The emerging role of glycolysis and immune evasion in gastric cancer[J]. Cancer Cell Int, 2023, 23(1): 317. doi: 10.1186/s12935-023-03169-1
[7] 翟冬妍, 吕冠华, 范颖. 旋覆代赭汤现代临床应用概述[J]. 中医药临床杂志, 2024, 36(2): 382-386. [8] 田晶晶, 袁红霞, 张月林, 等. 基于Caspase-3/Bcl-2/Bax信号通路探究加味旋覆代赭汤治疗食管癌前病变的作用机制[J]. 中国中西医结合外科杂志, 2024, 30(2): 258-264. doi: 10.3969/j.issn.1007-6948.2024.02.022 [9] 马媛, 荀敬, 王波涛, 等. 旋覆代赭汤对食管癌细胞干性的影响[J]. 中国应用生理学杂志, 2022, 38(1): 62-67. [10] 姚一菲, 孙可欣, 郑荣寿. 《2022全球癌症统计报告》解读: 中国与全球对比[J]. 中国普外基础与临床杂志, 2024, 31(7): 769-780. [11] TAKEUCHI H, MIYATA H, GOTOH M, et al. A risk model for esophagectomy using data of 5354 patients included in a Japanese nationwide web-based database[J]. Ann Surg, 2014, 260(2): 259-266. doi: 10.1097/SLA.0000000000000644
[12] ELLIOTT J A, DOCHERTY N G, ECKHARDT H G, et al. Weight loss, satiety, and the postprandial gut hormone response after esophagectomy: a prospective study[J]. Ann Surg, 2017, 266(1): 82-90. doi: 10.1097/SLA.0000000000001918
[13] 孙千惠, 朱广辉, 许博文, 等. 基于气陷理论探析恶性肿瘤术后中医药治疗与康复思路[J]. 中医杂志, 2024, 65(11): 1120-1125. [14] 李奕, 庞博, 花宝金, 等. 胃肠恶性肿瘤从"通道"立论的中医药治疗策略与思考[J]. 中华中医药杂志, 2023, 38(11): 5344-5347. [15] 田硕, 刘万里, 苏坤涵, 等. 旋覆代赭汤临床应用研究进展[J]. 亚太传统医药, 2021, 17(7): 205-209. [16] 邹利艳. 旋覆代赭汤联合针刺治疗胃癌术后胃瘫综合征31例[J]. 湖南中医杂志, 2020, 36(4): 52-54. [17] 向梅, 赵东丽, 梁宪斌, 等. 旋覆代赭汤联合耳穴按压对黑色素瘤化疗后不良反应的临床观察[J]. 实用中医内科杂志, 2020, 34(5): 109-112. [18] 谭林深, 朴瑛, 郑振东. 旋覆代赭汤加味治疗胰腺癌术后外分泌功能不全临床疗效观察[J]. 临床军医杂志, 2020, 48(1): 44-46. -
期刊类型引用(5)
1. 郭守刚. 水通道蛋白4抗体阳性的视神经脊髓炎谱系疾病的防复发治疗. 重庆医科大学学报. 2024(05): 593-596 .  百度学术
百度学术
2. 程畅,王伟,徐沙贝. MOG和AQP4抗体相关脱髓鞘疾病的研究进展. 神经损伤与功能重建. 2023(02): 96-99 . 百度学术
3. 代淑华,滕勇士,蒋玙姝,秦灵芝,薛红飞,徐佳佳,李玮. 视神经脊髓炎谱系疾病复发危险因素的研究进展. 实用临床医药杂志. 2023(08): 126-129+134 . 本站查看
4. 杨艳琼,周辉,陈东林,曾燕兰,罗利. 一例误诊为急性脑梗死的视神经脊髓炎谱系疾病及文献回顾复习. 智慧健康. 2023(10): 105-108+113 . 百度学术
5. 简佳芳,赵晓漪,何钱昆,吴远华. 吴远华治疗视神经脊髓炎临证经验. 贵州中医药大学学报. 2023(05): 41-45 . 百度学术
其他类型引用(2)
计量
- 文章访问数: 52
- HTML全文浏览量: 8
- PDF下载量: 5
- 被引次数: 7
 苏公网安备 32100302010246号
苏公网安备 32100302010246号
